
Evolution has always been portrayed as a forward march toward greater complexity and specialization. Yet beneath this familiar narrative lies one of biology’s most intriguing questions: can the evolutionary process actually turn around and head backward? Picture a snake suddenly sprouting legs again, or a bird redeveloping teeth after millions of years without them. While it sounds like science fiction, researchers are discovering that evolution might be far more reversible than we once thought.
The debate has intensified as new evidence emerges from laboratories around the world. Some studies show remarkable reversals in bacteria and simple organisms, while others reveal the seemingly insurmountable barriers that prevent complex features from returning. The implications stretch far beyond academic curiosity, touching on everything from antibiotic resistance to understanding our own evolutionary future.
The Origins of Scientific Doubt About Evolutionary Reversals

Belgian paleontologist Louis Dollo proposed in 1893 that “an organism never returns exactly to a former state, even if it finds itself placed in conditions of existence identical to those in which it has previously lived … it always keeps some trace of the intermediate stages through which it has passed.” This principle became known as Dollo’s law of irreversibility. The law wasn’t just an abstract concept but emerged from Dollo’s careful observations of fossil creatures that seemed to have lost complex features forever.
Dollo’s law gained traction because it aligned with common sense observations. According to Richard Dawkins, the law is “really just a statement about the statistical improbability of following exactly the same evolutionary trajectory twice (or, indeed, any particular trajectory), in either direction”. The reasoning seemed sound: once you’ve dismantled something complex, the chances of rebuilding it exactly the same way are astronomically small.
Stephen Jay Gould suggested that irreversibility forecloses certain evolutionary pathways once broad forms have emerged: “once you adopt the ordinary body plan of a reptile, hundreds of options are forever closed, and future possibilities must unfold within the limits of inherited design.” This perspective dominated evolutionary thinking for over a century.
The Molecular Machinery Behind Evolutionary Constraints
Modern genetics has revealed why reversing evolution might be so difficult. The culprit is something called epistasis, which sounds technical but describes a simple concept: the sign of the fitness effect of a mutation is under epistatic control; thus, such a mutation is beneficial on some genetic backgrounds and deleterious on others. Think of it like renovating a house where moving one piece of furniture affects where everything else can go.
Specific epistasis is caused by direct and indirect physical interactions between mutations, which nonadditively change the protein’s physical properties, such as conformation, stability, or affinity for ligands. In contrast, nonspecific epistasis describes mutations that modify the effect of many others. These interactions create a tangled web where undoing one change might require undoing several others first.
The implications are staggering. Extensive epistasis prevents reversions and necessitates fixation of new mutations, leading to a functionally identical sequence. Many amino acid exchanges between the new and original enzyme are not tolerated, implying sequence incompatibility. It’s like trying to unscramble an egg – theoretically possible, but practically impossible.
When Bacteria Defy the Rules
Recent experiments with bacteria have shown that simple evolutionary changes can indeed be reversed under the right conditions. MIT researchers studied whether bacteria could evolve resistance to cefotaxime but then lose it if they were placed in a new environment in which resistance to the original drug hindered their ability to survive. Genetic states that differ by only one mutation are always reversible if one state is more fit in one environment and the other is more fit in the other.
However, the reversibility window is narrow. “What we see in our system is that once the system gets four mutations, it’s unable to get back to where it started.” This finding suggests that evolution has a point of no return, at least for these bacterial systems. The door to reversal slams shut surprisingly quickly.
In laboratory studies, wild-type phosphotriesterase function could be restored with more than a 10,000-fold activity increase, but via an alternative set of mutations. The enzyme active site converged towards its original state, indicating evolutionary constraints imposed by catalytic requirements. Evolution found a way back, but it took a completely different route.
Complex Organisms Tell a Different Story
Regressive evolution involves the loss of previously evolved forms of complexity. An extreme example comes from the myxozoans, parasites with very simple anatomies – no mouths, nervous systems or guts – and very small genomes. These creatures represent evolution in seeming reverse, shedding complexity like autumn leaves. Yet they haven’t truly gone backward; they’ve adapted to their parasitic lifestyle in radical ways.
William R. Jeffery, a biologist at the University of Maryland, notes that “The chances that the same tape [of evolutionary changes] would be … reversed in the same way is highly improbable.” Evolution doesn’t really go backward in the sense of retracing evolutionary steps, experts say. What looks like reversal is often something entirely different.
Losses in complexity may accompany less-obvious increases in complexity, such as the biochemistries parasites use to get inside hosts. “It’s very easy for people … to think of evolution in terms of what you see … what the morphological features are,” but “there are also lots of other features that we don’t see at the physiological and the biochemical level.”
The Curious Case of Vanishing and Returning Features
Examples of evolutionary reversals include the re-expression of digits in horses and other animals long after their initial loss. The evolution of multicellularity in yeast and its subsequent reversal to unicellularity also exemplifies evolutionary reversal, highlighting the role of selective pressures in driving these changes. These cases challenge our assumptions about what can and cannot be undone in evolution.
In insects like phasmids, wings have been lost and regained multiple times, indicating that complex traits can indeed be reversible under certain conditions. Penguin ancestors were able to fly, but they lost that ability quickly after the demise of the large dinosaurs. The oldest known penguin, with fossils dating to around 60 million years ago, was already a flightless swimmer with stubby wings. These examples show that major transitions can happen relatively rapidly.
Experimental evolution studies in yeast have shown that multicellularity can evolve rapidly under selection, and when selection is reversed, unicellularity can re-evolve. This demonstrates that even major organizational changes can be reversed under the right selective pressures, although the genetic pathways involved in the reversal may differ from those in the original evolution.
The Hidden Genetic Archives
Cases where animals such as horses, guinea pigs, canids, and cetaceans have re-expressed digits or limb elements millions of generations after their loss are considered examples of atavisms, where suppressed regulatory genes are reactivated, leading to the reappearance of ancestral traits. This suggests that evolution doesn’t always completely erase the genetic instructions for lost features.
Large genomics databases and evo-devo studies are showing how the underlying developmental pathways and genetic architecture can be retained after the loss of a character. It’s like keeping the blueprint even after tearing down the building. Sometimes, under the right circumstances, construction can begin again using the old plans.
Reversals can occur through the reactivation of suppressed genes or through hybridization with lineages that retain the ancestral trait. However, not all apparent reversals are true evolutionary reversals; some may result from developmental anomalies or gene knockouts that mimic ancestral conditions without true evolutionary reversion. The distinction between genuine reversal and mere resemblance becomes crucial.
When Eyes See Again After Eons of Darkness

Living hagfish are remarkably unchanged from their ancient counterparts, and so scientists long thought that modern, sightless hagfish eyes represented a kind of intermediate step between primitive light-sensing spots and camera-like eyes. But when Sarah Gabbott trained an electron microscope on ancient fish fossils, she found remnants of pigment-containing structures within their eyes that would have allowed the animal to discern shapes and images. Ancient hagfish could see, but their descendants cannot.
Researchers continue to debate whether energy consumption or random mutations drive vision loss, with both processes likely working simultaneously. The loss of sight in cave-dwelling creatures has been observed repeatedly, but genuine restoration of complex vision remains elusive. “But the eye didn’t go in reverse. It just stopped going forward.”
The evolution of vision represents one of the most complex biological achievements. Once this intricate system breaks down, the chances of reassembling all its components in the right order become vanishingly small. Each step requires precise coordination between multiple genes and developmental pathways.
The Temperature of Change: Environmental Pressures
To evaluate the importance of selection for evolutionary reversals to unicellularity, researchers exposed different multicellular isolates to varying periods of UV radiation. Of the possible stresses often encountered by microorganisms, they selected UV radiation because it is easy to manipulate, imposes strong selection, and isolates vary in their response to this environmental factor. Environmental pressure can indeed drive reversal, but the results aren’t always predictable.
Although all populations converge to unicellular phenotypes the mode and tempo of evolution differs between populations and even within replicate lines of the same population. Even under identical conditions, evolution doesn’t follow the same path twice. The element of chance plays a larger role than scientists once suspected.
Selection, chance, and history are all likely to contribute to the reversibility of multicellularity, but their relative importance is still debated and might change during the transition itself. Strong selective pressures and high population sizes could overwrite the role of history and chance. Even in these cases, history and chance might be important for the dynamics of these transitions, shaping the tempo and mode of reversibility.
Modern Tools Reveal Ancient Secrets

Recent studies show that there are very likely some cases of ‘breaking Dollo’s law.’ As post-genomic research grows from showing patterns to revealing processes, new emphasis is needed on the molecular mechanisms by which Dollo’s law might be broken. Shifting the argument from ‘if it happened’ to ‘how it happened’ will provide richer understanding of organisational and evolutionary biology. Technology is transforming our understanding of what’s possible in evolution.
Using up to 194,358 SNP loci and 1,334,760 bp of sequence, researchers have reconstructed the phylogeny of viviparous and oviparous lineages of common lizards and inferred the evolutionary history of parity modes. They find strong statistical support for a topological arrangement that suggests a reversal to oviparity from viviparity. These genomic tools reveal evolutionary reversals that would have been impossible to detect just decades ago.
By genomic and molecular approaches, the depth of biological analyses possible are much richer today than ever before. Now it is not just the pattern but the mechanism that can be examined for the first time. This deep exploration is extremely fertile ground for evolutionary biologists seeking to understand the basis of adaptation to environment and the mechanisms of complex trait evolution.
Breaking the Law: Modern Challenges to Dollo’s Principle
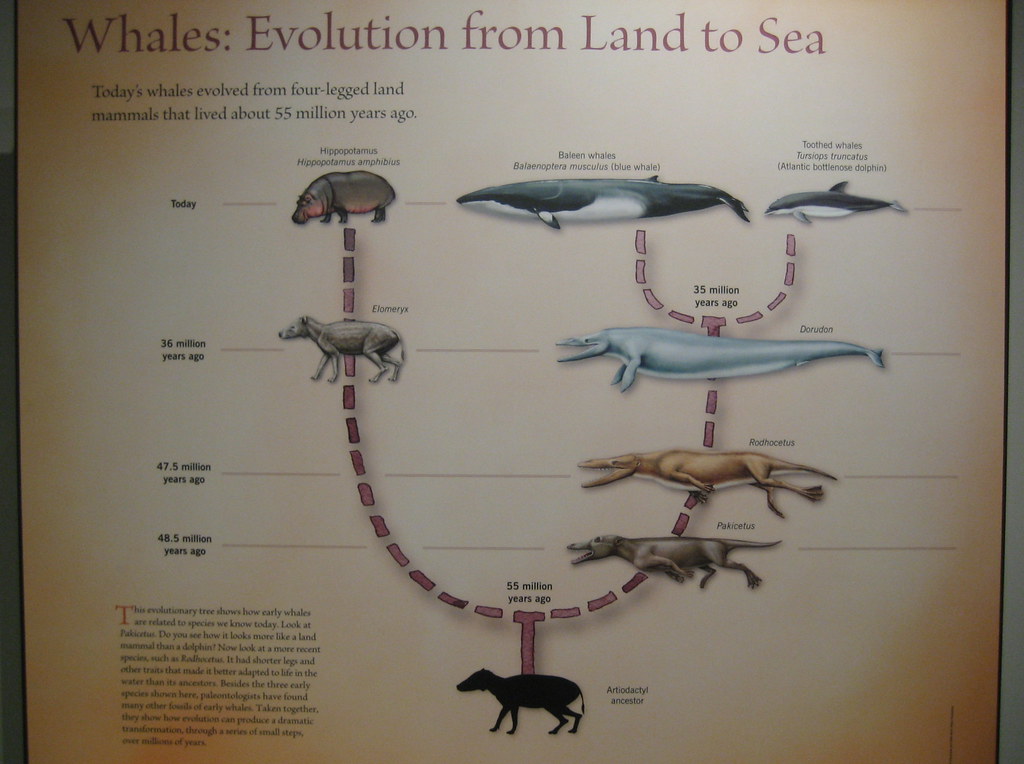
Dollo’s law has since been refuted by evidence that evolutionary specialisation can be undone. For instance, reversible evolution has been observed on a relatively short evolutionary timescale in the peppered moth. In the 19th century a dark morph of the moth emerged in response to air pollution during the Industrial Revolution and became the dominant colour morph, almost completely replacing the light-coloured form. By the late 20th century, however, the light morph was on the rise again, its increase coincident with the decline of air pollution in England.
Phylogenetic studies have recently demonstrated cases where seemingly complex features such as digits and wings have been reacquired. Some examples show that evolutionary losses of complex traits can be reversed, for instance, the regaining of the second molar in the lynx and the fifth premolar in an anthracotheriid. The number of nipples in mammals is extremely variable among species, indicating that reversals of changes are common. Other examples are the regaining of phalanges in several squamate taxa.
These discoveries have forced scientists to reconsider the absolute nature of evolutionary constraints. What once seemed like immutable laws now appear more like strong tendencies that can occasionally be overcome under the right circumstances.
Evolution’s capacity for reversal remains one of science’s most captivating mysteries. While simple changes can sometimes be undone, complex adaptations face nearly insurmountable barriers created by the tangled web of genetic interactions. The debate continues as new technologies reveal previously hidden evolutionary pathways, challenging our fundamental assumptions about the nature of biological change. Perhaps the most profound insight is that evolution, like life itself, refuses to be confined by our neat categories and predictions. What do you think about evolution’s potential for reversal? Tell us in the comments.


